
لمحة عامة عن بروتيز الاسبارتك من التصنيف الى التوصيف
صادق فنجان حسناوي1،2، زينة كاظم اليونس1، علاء جبار عبد المنهل1
1 قسم علوم الأغذية، كلية الزراعة، جامعة البصرة، البصرة 61004، العراق.
2 قسم الإنتاج الحيواني، كلية الزراعة، جامعة ميسان، ميسان 62004، العراق.
بريد الكتروني: pgsagr.sadeq.hasnawi@uobasrah.edu.iq ; zena.issa@uobasrah.edu.iq ; alaa.abd@uobasrah.edu.iq
HNSJ, 2024, 5(4); https://doi.org/10.53796/hnsj54/3
تاريخ النشر: 01/04/2024م تاريخ القبول: 08/03/2024م
المستخلص
يتزايد الطلب على الإنزيمات في القطاعات الصناعية بسرعة بسبب مزاياها الاقتصادية والبيئية ، أذ تنتج الكائنات الحية الدقيقة أنواعًا مختلفة من الإنزيمات ومن بين هذه الانزيمات هي انزيمات البروتيزات المنتجة من مصادر ميكروبية نتيجة لأنخفاض التكلفة وارتفاع معدل الإنتاج والتي اكتسبت اهتمامًا خاصًا في القطاعات الصناعية ، في هذه المراجعة قمنا بتلخيص المعلومات حول تصنيف البروتيزات اعتمادا على الاحماض الامينية الموجودة في الموقع الفعال او على اساس متطلباتها المعدنية أذ قسمت البروتيزات الى اربع مجاميع وهي بروتيزات الاسبارتك aspartic والسيرين serine والسستين cystein والثريونين threonine ، كما قسمت البروتيزات حسب الرقم الهيدروجيني الى ثلاثة اقسام هي البروتيزات الحامضية والقاعدية والمتعادلة ، وكذلك تحديد أهم مصادر انتاج البروتيزات ودراسة تركيب والية عمل بروتيز الاسبارتك والعوامل المؤثرة في انتاج البروتيزات من الفطريات وتنقية وتوصيف بروتيز الاسبارتك.
الكلمات المفتاحية: بروتيز الاسبارتك، البروتيزات الحامضية درجة الحموضة، ودرجة الحرارة، المادة الخاضعة
An overview of aspartic protease from classification to characterization
Sadeq Fenjan Hasnawi1,2, Zena Kadhim AL-Younis1, Alaa Jabbar Abd Al-Manhel 1
1Department of Food Science, College of Agriculture, University of Basrah, Basrah 61004, Iraq;
2Department of Animal Production, College of Agriculture, University of Misan, Misan 62004, Iraq
pgsagr.sadeq.hasnawi@uobasrah.edu.iq ; zena.issa@uobasrah.edu.iq ; alaa.abd@uobasrah.edu.iq
HNSJ, 2024, 5(4); https://doi.org/10.53796/hnsj54/3
Published at 01/04/2024 Accepted at 08/03/2024
Abstract
The demand for enzymes in industrial sectors is increasing rapidly due to their economic and environmental advantages , Microorganisms produce different types of enzymes, and among these enzymes are protease enzymes produced from microbial sources as a result of the low cost and high production rate, which have gained special interest in the industrial sectors ,In this review, we summarized the information about the classification of proteases based on the amino acids present in the active site or on the basis of their mineral requirements. The proteases were divided into four groups, which are aspartic proteases, serine, cysteine, and threonine. Proteases were also divided according to pH into three sections: proteases. Acidic, basic and neutral , As well as identifying the most important sources of protease production, studying the structure and mechanism of action of aspartic protease, the factors affecting the production of protease from fungi, and purifying and characterizing aspartic protease.
المقدمة
تعُد الإنزيمات محفزات حيوية تشارك تقريباً في جميع التفاعلات البيولوجية داخل الأنظمة الحية ، والتي استعملت في صناعة البيرة ، الخل و الجبن منذ عصور ما قبل التاريخ ، وأنتجت الانزيمات من سلالات مختارة من الكائنات الحية الدقيقة على نطاق واسع واستعملت طرق متطور في تنقيتها (Sharma, 2019) . وهذا أسهم بشكل كبير في تصحيح العمليات الصناعية واحدث ثورة في تطوير التكنولوجيا الحيوية نظرًا لكفاءتها وفعاليتها كمحفزات عند اقل تركيز(Gimenes et al., 2021) .
استعملت العديد من الإنزيمات في الصناعات المختلفة مثل اللايبيز ، والبروتيز ، والأميليز ، والسليوليز ، والزايليز وغيرها ، ومع ذلك ، تبقى انزيمات البروتيز هي النوع السائد لأنها ذات قيمة كبيرة من خلال تطبيقاتها الواسعة في مختلف المجالات منها منتجات الألبان والأغذية والورق والمنظفات ، إذ تشكل البروتيزات حوالي 60 ٪ من إجمالي المبيعات للأنزيمات التجارية في كل عام (Pawar et al., 2023) وكما مبين في الشكل (1), اذ بلغ السوق العالمي للبروتيزات في عام 2019 مايقارب 2.76 مليار دولار أمريكي ، ومن المتوقع أن يزداد خلال الفترة من 2019-2024 بمعدل نمو سنوي قدره 6.1٪ (Choudhary et al., 2022) .
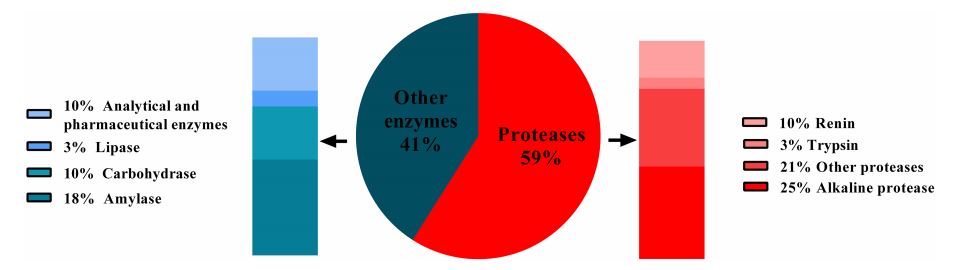
الشكل (1) المبيعات السنوية للأنزيمات (Pawar et al., 2023)
ان البروتيزات او ما تسمى ببتيدات التحلل المائي، هي انزيمات لها القدرة على كسر الروابط الببتيدية في البروتينات (Koistinen et al., 2023) . وهي تلعب دور مهم في العديد من العمليات البيولوجية (Couture, 2023) ، بما في ذلك الهضم (Kalaiselvan et al., 2023) ، تخثر الدم (Dasari et al., 2023) ، الاستجابة المناعية (Dudani et al., 2018) ، وتشارك أيضا في تنظيم نمو الخلايا فضلاً عن العديد من الوظائف المهمة للكائنات الحية (Erekat, 2022; Zhang et al., 2023). توجد هذه الانزيمات في جميع الكائنات الحية ، مثل الحيوانات، النباتات، البكتريا والفطريات (Naveed et al., 2021) . تعمل البروتيزات على الاصرة الببتيدية الموجودة في البروتينات وتحللها مائيا Proteolysis enzyme الى ببتيدات واحماض امينية، وبصورة عامه فانها اما ان تكون بروتيزات حامضية اوقاعدية اومتعادلة, كما تعتبر البروتيزات الحامضية من اهم البروتييزات المستعملة في التصنيع الغذائي (Patil et al.,2012; Al-Manhel.,2017 ). لذا فان الهدف من هذه الدراسة هو معرفة تصنيف ومصادر البروتيزات كذلك دراسة تركيب والية عمل بروتيز الاسبارتك لندرة الدراسات التي تناولت هذا الانزيم بشكل مفصل على الرغم من اهميته وتطبيقاته الواسعة فضلا عن توفير فهم أفضل للعوامل المختلفة المؤثرة في انتاج البروتيزات بشكل عام من الفطريات .
تصنيف البروتيزات Classification of protease
وفقًا للجنة التسمية التابعة للاتحاد الدولي للكيمياء الحيوية والبيولوجيا الجزيئية International Union of Biochemistry and Molecular Biology (IUBMB) ، تم تصنيف البروتيزات ضمن المجموعة الثالثة المتمثلة بأنزيمات التحلل المائي hydrolases ، كما قسمت البروتيزات تبعا لفعلها التخصصي بحيث تكون اما داخليه Endopeptidases او خارجية Exopeptidases (Pawar et al., 2023) .
الببتايديزات الخارجية Exopeptidases
تعمل الببتايديزات الخارجية بالقرب من النهايات الطرفية في السلسلة الببتيدية بناءاً على موقع عملها والتي تقوم بتحليل جزيئة البروتين من خلال تكسير الأواصر الببتيدية الطرفية محررة بالتعاقب الأحماض الأمينية الحرة إلى وسط التفاعل، تقسم الببتايديزات الخارجية الى نوعين اعتماداً على موقع عملها في السلسلة الببتيدية الأولى تسمى الببتايديزات الأمينية Amino Peptidase والتي تعمل على النهاية الطرفية N في السلسلة الببتيدية، اما الثانية تدعى الببتايديزات الكاربوكسيلية Carboxyl Peptidase والتي تعمل على النهاية الطرفية C في السلسلة البببتيدية (Mienda et al.,2014) .
الببتايديزات الداخلية Endopeptidases
تعمل الببتايديزات الداخلية على الاواصر الببتيدية في المواقع الداخلية من السلسلة الببتيدية بعيداً عن النهايات الطرفية N و C (Mienda et al.,2014) والتي عندها يسمى الانزيم (proteinase)، هذا النوع من البروتيزات (EC 3.4.11-24) يقسم الى اربع مجاميع وهي بروتيزات الاسبارتك aspartic والسيرين serine والسستين cystein والثريونين threonine اعتمادا على الاحماض الامينية الموجودة في الموقع الفعال او على اساس متطلباتها المعدنية (Mamo and Assefa., 2018 , Rawlings and Salvesen.,2013) .
- بروتييزات الاسبارتك Aspartic protease (EC 3.4.23)
ينتمي بروتيز الاسبارتك الى مجموعة من البروتيزات التي تعتمد على النشاط التحفيزي لوحدتين من حامض الاسبارتك (Gurumallesh et al., 2019). ويطلق على هذا النوع من البروتيزات بالبروتيزات الحامضية Acid Proteases او البروتيزات الكاربوكسيلية Carboxylic Proteases (Majid, 2017) والتي تعمل في وسط حامضي على تحليل الاصرة الببتيدية التي تربط بين الاحماض الامينية، اذ تحتوي هذه الأنزيمات على حامض الاسبارتيك والتايروسين في موقعها الفعال ويكون الرقم الهيدروجيني الأمثل لفعاليتها ( 3-5 ) والوزن الجزيئي يتراوح بين ( 30 – 45 ) كيلودالتون ودرجة الحرارة المثلى (40-55 ) مْ ( (Gimenes et al.,2021 ; Mamo and Assefa, 2018 ، واستعملت البروتيزات الحامضية في ازالة العكارة من البيرة وعصير الفاكهة وايضاً في تطرية اللحوم وكذلك في معالجة حالات عسر الهضم ، تتواجد في الحيوانات والنباتات والفطريات والفايروسات ، غالباً ما تنتج خارج الخلية عن طريق الانواع الفطرية مثل Aspergillus niger ، Aspergillus oryzae ، Aspergillus awamori ، Aspergillus fumigates ، Aspergillus saitoi ونادراً ما توجد في البكتريا والابتدائيات (Wei et al.,2023)، وهي غير حساسة لمثبطات الأنزيمات الأخرى، الا انها تثبط تماماً عند وجود مادة الببستاتين pepstatin Mamo et al., 2020) ).
- البروتييزات المعدنية Metallo protease ( EC 3.4.24)
تتطلب هذه المجموعة من الإنزيمات أيون فلز ثنائي التكافؤ لعملها ، مثل الزنك أو الكوبالت أو المنغنيز لذلك تسمى بالبروتيزات الفلزية، وهي بهذا تكون حساسة للعوامل المخلبية مثل (EDTA ) Ethylene diamine tetra acetic acid والتي تعمل على ازالة الايون المعدني من الانزيم مما يؤدي الى تثبيطه Singh et al.,2016) ) ، تحتوي هذه الأنزيمات على الحامض الاميني فينيل الآنين أو ليوسين في موقعها الفعال ويكون الرقم الهيدروجيني الأمثل لفعاليتها ( 5-7) والوزن الجزيئي يتراوح بين ( 19-37 ) كيلودالتون ودرجة الحرارة المثلى (65-85 ) مْ ( (Mamo and Assefa, 2018 ، ومن انزيمات هذه المجموعة هو انزيم Thermolysin المنتج من بكتريا Bacillus thermoprotoolyticus والذي يحتوي على ايون الزنك Zn+2 في موقعه الفعال (Razzaq et al.,2019) .
- بروتييزات السيرين Serein protease) )( EC 3.4.21)
تحوي هذه الأنزيمات على الحامض الاميني ( السيرين أو الهستدين أو الأسبارتك ) في الموقع الفعال (Majid, 2017) وتثبط عند وجود مركب EDTA و triamino acetic acid وهي بروتيزات قاعدية يكون رقمها الهيدروجيني الأمثل لفعاليتها بين (6-11) اما الوزن الجزيئي له فيتراوح من ( 18 – 35 ) كيلودالتون ودرجة الحرارة المثلى لعملها بين (50-70 ) مْ ( (Mamo and Assefa, 2018 ، تعد بروتيزات السيرين من اهم البروتيزات التي تم فصلها وتنقيتها من الاجزاء النباتية المختلفة (Majid, 2017) ، في حين ان البكتريا هي الاكثر شيوعاً في انتاج هذا النوع من البروتيزات مقارنة مع الاحياء المجهرية الاخرى ومن أهم الاجناس البكتيرية السائدة هي Pseudoalteromonas, Psychrobacter photobacterium , Vibrio , halobacillus , Bacillus , Microbulbifer , and shewanella. تعود بعضها الى الانواع Vibrio etschnikovii ، Bacillus sp. CEMB10370 ، Pseudomonas sp. ، Bacillus alcalophilus LW8 بينما انتجت هذه الإنزيمات من انواع محددة من الاعفان مثل عفن Aspergillus tamari ) Banerjee & Ray, 2017 ( .
- بروتييزات السستين (Cysteine protease) ( EC 3.4.22)
تحتوي هذه الأنزيمات على الحامض الاميني السستين والهستدين في الموقع الفعال ( (Mamo and Assefa, 2018 وتسمى أيضا بروتيزات الثايول Thiolاو بروتيزات السلفهايدرل Sulfhydryl وهي حساسة لمركبات السلفهيدريل (sulfhydryl reagent) مثل iodoacetamide وiodacetic acid وTLCK والفلزات الثقيلة، وتنشط بالعوامل المختزلة كالمواد المخلبية والسستائين وسيانيد البوتاسيوم (chelating agent) مثل EDTA (Majid, 2017)، وتكون هذه الانزيمات نشطة عند درجة الحموضة المتعادلة وتقع الفعالية المثلى لمعظم هذه الأنزيمات ضمن مدى من الأرقام الهيدروجينية التي تتراوح بين (5- 8) ووزنها الجزيئي (34-35) كيلودالتون ودرجة الحرارة المثلى (40-55) مْ ( (Mamo and Assefa, 2018، انتجت من بعض الاجناس البكتيرية مثل Bacillus spp. (Razzaq et al.,2019)، كما وجدت هذه الانزيمات في مجموعة من النباتات تحت اسم الباباين Papain والبروملين bromelain والكيموباباين Chymopapin والفيسين ficin والكاريكين Caricain أيضا في الفيروسات والطفيليات والحشرات (Majid, 2017). وبصورة عامة يبين جدول (1) خواص وصفات بروتيزات الاسبارتك والسستين والمعدنية والسيرين .
جدول (1) خواص انواع البروتيزات الاربعة من مصادر مختلفة
(;Majid.,2017 Gurumallesh et al.;2019 ( Mamo and Assefa, 2018 ;
| البروتيز | EC | الوزن الجزيئي
KDa |
pH
الامثل |
درجة الحرارة
المثلى |
الحامض الاميني الفعال | المثبط | مصدر البروتيز |
| بروتيز السيرين | 3.4.21 | 18-35 | 6-11 | 50-70 | السيرين او الهستدين او الاسبارتك | EDTA ,
triamino acetic acid |
Bacillus, Aspergillus, Animal tissue (gut), Tritirachium album (thermostable) |
| بروتيز السستين او الثايول | 3.4.22 | 34-35 | 5-8 | 40-55 | السستين والهستدين | Iodoacetamide,
p-CMB |
Aspergillus, stem of pineapple (Ananas comosus), latex of fig tree (Ficus sp), papaya (Carica papaya), Streptococcus, Clostridium |
| بروتيز الاسبارتك
او الكاربوكسيل |
3.4.23 | 30-45 | 2-5 | 40-55 | الاسبارتك ،
الاسبارتك |
Pepstatin | Aspergillus, Mucor, Endothia, Rhizopus, Pencillium, Neurospora, animal tissue (stomach) |
| البروتيز المعدني | 3.4.24 | 19-37 | 5-7 | 65-85 | فينل النين او ليوسين | EDTA and
EGTA |
Bacillus,
Aspergillus, Pencillium, Pseudomonas, Streptomyces |
التصنيف حسب الرقم الهيدروجيني
قسمت البروتيزات حسب الرقم الهيدروجيني الى ثلاثة اقسام هي: (Gimenes et al., 2021 , Solanki et al.,2021):
- البروتيزات الحامضية Acid proteases :
وتشمل البروتيزات المنتجة من الحيوانات مثل الرنين renin و الببسين pepsin ، والبروتيزات المنتجة من النباتات مثل البابين papain والبروملين bromelain والفيسين ficin ، تعمل هذه البروتيزات عند رقم هيدروجيني ( 3 – 5.6 ) (Machado et al., 2016; Razzaq et al., 2019) . تعُد الفطريات المصدر الرئيسي لأنتاج البروتيز الحامضي ، وان الاجناس الاكثر شيوعاً في انتاجه هي Aspergillus وPenicillium وEndothia وMucor (Mammo and Assefa, 2018). هناك العديد من الابحاث التي أكدت على اهمية انتاج البروتيزات الحامضية بواسطة الاعفان (Mandujano-González et al. 2016) . اذ انتج da Silva et al. (2016) بروتيز الاسبارتك من عفن Rhizomucor miehei، والذي يمكن أن يكون بمثابة إنزيم بديل لأنزيم تخثر الحليب أثناء إنتاج الجبن، كما لاحظ (Souza et al. (2017 عند توصيف البروتيز الحامضي المنتج من عفن Aspergillus foetidus ان الرقم الهيدروجيني الامثل لنشاط البروتيز (5 ) عند درجة حرارة مثلى (55 ) مْ .
2- البروتيزات المتعادلة Neutral proteases :
وهي البروتيزات المنتجة من البكتريا والفطريات والتي تعمل عند رقم هيدروجيني (7 ) ، وتشمل البروتيزات المعدنية التي تتطلب اثناء نشاطها وجود ايون موجب الشحنة ثنائي التكافىء (Contesini et al. 2017; Razzaq et al. 2019). استعملت البروتيزات المتعادلة لأغراض مختلفة منها صناعة الخبز والاجبان والادوية والمنظفات وغيرها (Razzaq et al. 2019) . وقد عمل Ao et al. (2018) على انتاج وتوصيف البروتيز المتعادل من عفن Aspergillus oryzae Y1 المعزول من الفاصوليا العريضة المخمرة ، اذ لاحظ امتلاكه وزن جزيئي ( 45 ) كيلودالتون ورقم هيدروجيني (7) ودرجة حرارة مثلى ( 55) مْ . كما اشار (De Oliveira et al. (2020 عند إنتاج البروتيز المتعادل من عفن Moorella speciosa ان الرقم الهيدروجيني الامثل لعمل الانزيم هو ( 7) .
- البروتيزات القاعدية Alkaline proteases :
تنتج هذه البروتيزات من الحيوانات والنباتات والبكتريا والفطريات والتي تعمل عند رقم هيدروجيني ( 8 – 12 ) ومن أهم انواع البروتيزات القاعدية هي subtilisin ، Carlsberg و subtilisin novoوألتي أنتجت من بكتريا Bacillus (Contesini et al. 2017) . إذ إن البروتيزات القاعدية المنتجة من جنس B. subtilis والمستخدمة للأغراض التجارية تعد أمنة (Zhang et al. 2017) . هناك العديد من الأبحاث التي تناولت إنتاج البروتيزات القاعدية من جنس Bacillus ، إذ قام Mothe and Sultanpuram (2016) بتنقية وتوصيف البروتيز القاعدي المنتج من بكتريا B. caseinilyticus . على الرغم من أن البروتيز القاعدي المنتج من جنس Bacillus هو الأكثر انتشارًا واستخدامًا، إلا أن هناك الكثير من الابحاث التي اشارت الى امكانية انتاج البروتيز القاعدي من انواع اخرى من الاحياء المجهرية مثل بكتريا Streptomyces sp. Solanki et al., 2021) ) . والفطريات (Sharma ., 2019)، اذ انتج ) da Silva et al. (2018 البروتيزات القاعدية من عفن Aspergillus tamarii URM4634 وكان افضل نشاطللبروتيز عند رقم هيدروجيني (8) .
تركيب وألية عمل انزيم بروتيز الاسبارتك
التركيب الاولي لأنزيم البروتيز المنتج من خميرة Candida tropicalis يتكون من 320 -340 حامض اميني ، والموقع الفعال في انزيم البروتيز يحتوي على وحدتين من حامض الاسبارتك احداهما تقع في الموقع 215 والاخر يقع في الموقع 32 ، اما التركيب الثانوي له فيتكون من شق عميق يحتوي على خمسة خيوط (sheet β-) محاطة بستة من التراكيب الحلزونية (helices – α ) كما مبين في الشكل ( 2 ) Theron.,2014 )) .
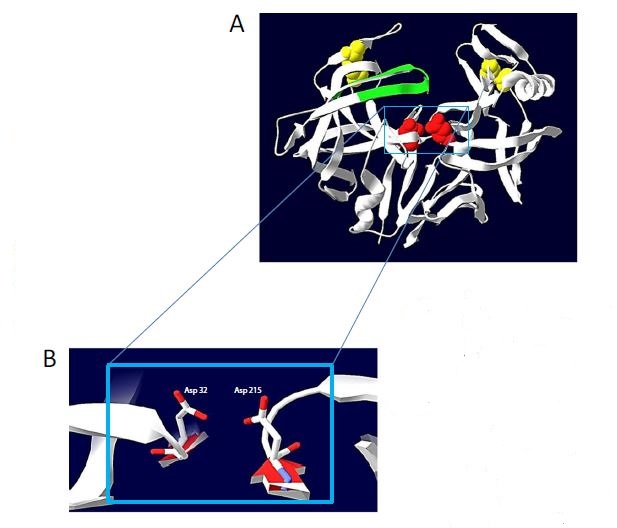
الشكل (2) تركيب انزيم البروتيز المنتج من خميرة Candida tropicalis Theron,2014 ) )
يُعد بروتيز الأسبارتيك من البروتيزات التي لها القدرة على كسر الروابط الببتيدية في البروتينات وتحللها مائيا الى ببتيدات واحماض امينية (Couture, 2023). اقُترح عدد من الآليات المختلفة لعمل الانزيمات وهي التحفيز ألحامضي _ القاعدي Acid – base catalysis ، التحفيز التساهمي Covalent catalysis ، التحفيز الايوني المعدني Metal ion catalysis ، التحفيز الالكتروستاتيكي Electrostatic catalysis ، الأ إن الطريقة الأكثر قبولًا على نطاق واسع لألية عمل بروتيز الاسبارتك هي آلية تحفيز القاعدة والحامض Acid-base catalysis إذ يتضمن التحفيز الحامضي قيام حامض الاسبارتك الموجود في الموقع الفعال بمنح البروتون للمادة الخاضعة ، اما في التحفيز القاعدي يعمل حامض الاسبارتيك الموجود في الموقع الفعال باستقبال البروتون من المادة الخاضعة وكما موضح في الشكل (3) ، إذ إن الموقع الفعال في الإنزيم يحتوي على وحدتين من حامض الاسبارتيك احدهما يقع في الموقع 32 من السلسلة الببتيدية الذي يعمل على منح البروتون للمادة الخاضعة ، بينما الاخر يقع في الموقع 215 من السلسلة الببتيدية، والذي يعمل على سحب البروتون من الماء وتكوين النيوكلوفيل السالب الشحنة ) (–oH مما يؤدي الى مهاجمة المادة الخاضعة وبالتالي كسر الرابطة بين النتروجين والكاربون وتكون الببتيد الاول ، اما حامض الاسبارتك الثاني الذي يقع في الموقع 32 من السلسلة الببتيدية يعمل على سحب البروتون من مجموعة الكاربونيل للجزء الاخر من المادة الخاضعة، وبذلك ينتج الببتيد الثاني للمادة الخاضعة (Brik and Wong , 2003 , Coates et al ,2008 , Wangsell ,2006) .
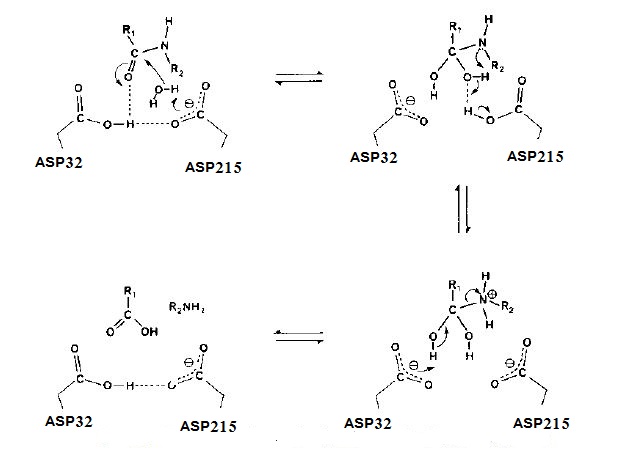
الشكل (3) الآلية المقترحة لعمل أنزيم بروتيز الأسبارتيك ( Coates et al.,2008 )
مصادر انتاج البروتيز Sources of protease production
تلعب البروتيزات دوراً مهماً في كل الكائنات الحية لذلك فهي توجد في مدى واسع من المصادر مثل الحيوانية والنباتية والميكروبية (Naveed et al., 2021) .
1- المصادر الحيوانية
تعُد انزيمات التربسين Trypsin ، الكيموتربسين Chemotrypsin ، الرنين Renein والببسين Pepsin من اكثر البروتيزات الحيوانية شيوعاً واستعمالاً في مجالات مختلفة ، إذ يعد التربسين الانزيم الهضمي المعوي المسؤول عن التحلل المائي لبروتينات الغذاء Singh et al.,2016) ). اما انزيم الرنين المستخلص من المعدة الرابعة للعجول الرضيعة فقد استعمل في صناعة الاجبان المختلفة (Gimenes et al., 2021). في حين استبدل انزيم الكيموتربسين المستخلص من منطقة البنكرياس في التطبيقات التشخيصية والتحليلية ، اما انزيم الببسين الذي يوجد في معدة جميع انواع الفقريات تقريباً فأنه ادخل في صناعة المنظفات N Jisha et al.,2013) ). ولقد لاحظ Dadshahi et al.,(2016) عند استخلاص وتنقية البروتيزات من الروبيان بانها ثابتة تجاه الحرارة العالية وتعمل عند الرقم الهيدروجيني ( 7-9 ) .
2- المصادر النباتية
استعملت البروتيزات النباتية على نطاق واسع في التطبيقات المختلفة مثل تطرية اللحوم وتخثر الحليب والهضم ولعلاج بعض الإمراض الفيروسية والسرطانية (Singh et al., 2016). تعُد انزيمات البروميلين والباباين من أبرز الإنزيمات النباتية المحللة للبروتينات ، وهما من عائلة الأناناس Bromeliaceae وCarica papaya على التوالي ، ومن اهم مصادر انتاج انزيم البروميلين هي سيقان الأناناس ولحاءه وأوراقه ،اما مصادر انتاج انزيم الباباين فهي نبات البابايا (Gimenes et al., 2021) . ومن البروتيزات المهمة ايضاً هي انزيمات الكيراتينيز Keratinase، التي استعملت في تحلل بروتينات الشعر والصوف وأنتاج الأحماض الأمينية الأساسية ، اكتسبت البروتيزات النباتية اهتماما متزايدا ، على الرغم من أن عملية إنتاج البروتيز من النباتات تستغرق وقتا طويلا أو قد تكون مكلفة من الناحية الأقتصادية Singh et al., 2016)). لاحظ (2016) Sun et al. عند تقييم ما يقارب من تسعين مصدراً نباتيًا؛ إن أعلى الإنزيمات المحللة للبروتين موجودة في الأناناس والتين والبابايا .
3- المصادر الميكروبية
تعُد الكائنات الحية الدقيقة من المصادر المهمة لأنتاج البروتيزات لما تتميز به من مزايا تتمثل في سرعة الانتاج ومساحة نموها المحدودة وبالتالي قلة التكاليف فضلاً عن سهولة التلاعب الجيني في تركيبها الوراثي وعدم تأثرها بالظروف المناخية (Bhatia et al. 2021 ; Putatunda et al. 2019). أما من الناحية التجارية ، فأن البروتيزات البكتيرية والفطرية قد حظيت بالكثير من الاهتمام Gurumallesh et al.;2019) ( ، اذ تشكل البروتيزات الميكروبية ثلثي إجمالي البروتيزات المستعملة في الصناعات المختلفة ، ومن اهم هذه البروتيزات هي البروتيز القاعدي البكتيري والبروتيز الحامضي الفطري ،اذ تم عزل الكائنات الحية الدقيقة المنتجة للبروتيز من مجموعة متنوعة من البيئات مثل الجذور والتربة مثل تربة المسالخ والصرف الصحي والمخلفات الصناعية والزراعية وما إلى ذلك (Jadhav et al. 2020; Hakim et al. 2018; Ash et al. 2018; Prajapati et al. 2017) .
تستعمل البروتيزات الميكروبيه في مدى واسع من التطبيقات المختلفة ، وتنتج بطرق مختلفة اعتماداً على نوع الكائن الحي المجهري المستعمل ولعل أفضل الطرق هي استعمال تقنية تخمرات الحالة الصلبة (Usman et al.,2021) وذلك لأنها عادة ما تكون أنزيماتها خارجية اي تفرز خارج خلايا الأحياء المجهرية etal.,2020) Mamo ) ، ولسهولة تنمية هذه الاحياء والتعامل معها وامكانية السيطرة على ظروف الانتاج وقصر مدته كذلك امكانية تسخير الامكانات المحلية لإنتاجه باقل التكاليف من خلال الاستفادة من المخلفات الزراعية والصناعية كالنخالة والكسبة وقشور الرز وغيرها (Novelli et al., 2016) .
تعُد الفطريات من الكائنات المجهرية التي تميزت في انتاجها للبروتيزات الحامضية (Yin et al.,2020;Patil et al.,2012 ) مقارنة مع الاحياء المجهرية الاخرى ولها خواص مشابهة للانزيمات الحيوانية ، والتي بدورها تقسم الى نوعين ,الاول هي البروتييزات الشبيهة بالببسين Pepsin like acid protease والتي تميز عفن Aspergillus (aspergillopepsin, EC 3.4.23.18) و Penicillium و Rhizopus ( rhizopuspepsin, EC 3.4.23.21 ) في انتاجها (Souza et al.,2017) اما النوع الثاني هي البروتييزات الشبيهه بانزيم الرنين Rennin like acid protease والتي انتجت من قبل عفن Mucor و Rhizomucor وRhizopus ( mucorpepsin, EC 3.4.23.23) ) وEndothia parasitica (endothiapepsin, EC 3.4.23.22) (Aljammas et al.,2018; Daudi et al.,2015;) تعد البروتيزات الحامضيه او ما يسمى ببروتيزات الاسبارتك proteases aspartic (EC 3.4.23 ) والتي تميزت الاعفان في انتاجها Mamo et al.,2020) ) الاكثر استعمالا في الصناعات الغذائية كصناعة الاجبان وتطريه اللحوم وتحضير المتحللات البروتينية وترويق العصائر وتكون فعاليتها القصوى وثباتيتها عندما يكون الرقم الهيدروجيني (5-2) الا انها تعمل عندا رقم هيدروجيني (6) ( Mamo and Assefa, 2018;Sun etal.,2018; Purushothaman etal.,2019;Yujie et al.,2019 ) .
العوامل المؤثرة في انتاج البروتيزات الحامضية ( الاسبارتك) من الفطريات
تلعب الظروف المثلى دوراً مهماً في إنتاج الإنزيمات أذ يشكل الوسط ألزرعي 30% من الكلفة الكلية لعملية التخمر وبالتالي فهو يعتبر عاملاُ مهما في نمو الاحياء المجهرية المنتجة للأنزيمات (dos et al., 2018) . ان التغيرات في درجة الحموضة، ودرجة الحرارة، والمادة الخاضعة ، ومحتوى الماء، وحجم اللقاح، وما إلى ذلك لها تأثير كبير على الأنشطة الأيضية للكائنات الحية الدقيقة, وتختلف هذه المعايير بشكل كبير من نوع إلى آخر، ولذلك فإن معرفة الظروف المثلى لنمو الفطريات لإنتاج معظم الإنزيمات يعد أمراً ضرورياً (Othman et al., 2023) . أذ ان حاجة الكائنات الحية لمصادر الكربون والنيتروجين تختلف من كائن حي إلى آخر، أو حتى بين نفس الأنواع المعزولة من مصادر مختلفة ( Hamza et al., 2023). الوسط الزرعي يجب ان يحتوي على مصدر كاربوني ومصدر نتروجيني ، فالمصدر الكاربوني يعتبر محفزاً لإنتاج الإنزيمات اومثبطا لها وهذا يعتمد على نوع المصدر الكاربوني وتركيزه في الوسط ، وكذلك نوع الكائن المجهري المستخدم للأنتاج ، كما ان المصدر النتروجيني يزود الوسط الزرعي اما بالأملاح اللاعضوية التي قد تكون مثبطاً لإنتاج الانزيم عند اضافتها للوسط مثل كلوريد الامونيومNH4Cl ،او تجهز الوسط الزرعي بالمصادر العضوية التي تحفز نمو الكائن المجهري مثل البروتين والاحماض الامينية وغيرها، وبالتالي فأنها تلعب دوراً كبيراً في تخليق الإنزيمات، هناك بعض الاملاح تضاف الى الوسط الزرعي بتراكيز معينة والتي تكون مصدراً للمعادن النادرة فضلاً عن الكبريت والفوسفات والكالسيوم مثل (CaCl2) للمحافظة على درجة الحموضة وبالتالي تحافظ على ثباتيه الانزيم (Aunstrup et al., 1979). اشار Othman et al. (2023) في دراسته لأنتاج انزيم البروتيز الحامضي من عزلة Aspergillus niger بأستعمال تخمرات الحالة الصلبة ، الى ان إضافة مصادر الكربون والنيتروجين إلى الوسط الصلب أدت إلى زيادة إنتاج الإنزيمات وتوليفها، كأستعمال الكازين كمصدر للنيتروجين والفركتوز كمصدر للكربون. كما لاحظ Bano et al. (2016) ارتفاع معدل إنتاج انزيم البروتيز عند تنمية فطر Pleurotus eryngii في وسط معدني يحتوي على 0.5% سكروز و1% كازين . ان الاختلاف في نسبة محلول الترطيب المستخدمة في تخمرات الحالة الصلبة يؤثر بشكل كبير على نمو وايض الاحياء المجهرية مما يؤثر سلباً وايجاباً على انتاجيتها لأنزيم بروتيز الاسبارتك ( Bensmail et al., 2015) ، اذ ان انخفاض وارتفاع نسبة محلول الترطيب يسبب قلة المغذيات وبالتالي يوفر بيئة غير ملائمة لنمو ونشاط الاحياء المجهرية أذ ان رطوبة المادة الصلبة يجب ان لا تقل عن 12 – 15 % والتي تعد الحد الأدنى لتوقف الفعاليات الحيوية للأحياء المجهرية (Luti et al., 2014). واشار Othman et al. (2023) في دراسته لأنتاج انزيم البروتيز الحامضي من عزلة Aspergillus niger بأستعمال تخمرات الحالة الصلبة ، الى ان افضل نسبة لمحلول الترطيب في انتاج الانزيم هي 1:0.75 .
كذلك تُعد الأنشطة الأيضية للكائنات الحية الدقيقة حساسة للغاية لتغيرات الرقم الهيدروجيني، كونه يؤثر على العديد من العمليات الأنزيمية ونقل العناصر الغذائية المختلفة عبر غشاء الخلية مما يضمن نمو وإنتاج المنتجات الايضية ، تؤدي هذه الأنشطة حتمًا إلى تحول أيضي في توازن أيونات الهيدروجين وبالتالي الرقم الهيدروجيني للوسط الزرعي (Othman et al. 2023) ، وان التباين في قيم الرقم الهيدروجيني يؤثر على ذائبية المغذيات وجاهزيتها في الوسط الزرعي وهذا ينعكس سلباً وإيجابا على سير العمليات الايضية وبالتالي تخليق وانتاج الانزيم (Whitaker , 1972) .
اوضح Bensmail et al. (2015) بأن اقصى إنتاج لأنزيم البروتيز بواسطة عفن A. niger FFB1 كان عند رقم هيدروجيني 4 ، في حين انخفضت الانتاجية عند جميع قيم الرقم الهيدروجيني الأعلى من 4 , في المقابل ذكر Chahrazad et al. (2021) ان درجة الحموضة المثلى لأنتاج انزيم البروتيز من عزلة Aspergillus niger كان عند )3 ( pH=، وأشارت النتائج إلى أن السلالة المختارة كان لها أيضًا نشاط تحلل بروتيني عند درجة حموضة محايدة، لكنه كان أقل أهمية، وقد يقترح أن السلالة أنتجت على الأقل اثنان من البروتيز، أحدهما حامضي بدرجة حموضة مثالية 3 والآخر قلوي عند درجة حموضة 7 . كما بين Mamo et al. (2020) ان الرقم الهيدروجيني الامثل لأنتاج انزيم البروتيز الحامضي من عزلة Aspergillus oryzae DRDFS13 هو (pH =5) . وكذلك تُعد درجة الحرارة من العوامل الاساسية المؤثرة بشكل كبير على معدلات نمو وانتاج الاحياء المجهرية اذ تختلف درجات الحرارة حسب نوع الكائن المجهري المستخدم (Banerjee et al., 2017) ، كما تؤثر على معظم التفاعلات الايضية للأحياء المجهرية بالأخص النشاطات الأنزيمية في الخلية من خلال تأثيرها على الصفات الفيزيائية للغشاء الخلوي, إذ بين (Othman et al . (2023 ان الحد الأقصى لإنتاج انزيم البروتيز الحامضي من عفن Aspergillus niger كان عند درجة حرارة 35 مْ الا ان الارتفاع عن هذه الدرجة يؤدي الى الانخفاض في انتاجية الانزيم حتى يفقد فعاليته عند 45 مْ بسبب إطلاق الحرارة الأيضية التي تسبب تجفيف الوسط الزرعي وبالتالي انخفاض النشاط المائي الذي يؤدي الى انخفاض توافر العناصر الغذائية، مما يقيد التهوية ويقلل من نمو وانتاج الاحياء المجهرية أو قد يكون ناتج عن دنترة البروتينات وتثبيط الإنزيم وانخفاض نمو الفطريات . كما لاحظ كلاً من (Khademi et al . (2013 و Foda et al . (2012) أن انتاج البروتيز من A. niger باستخدام تخمرات الحالة الصلبة يحدث بشكل أفضل عند درجة الحرارة (30) مْ, اشار (2020) Chimbekujwo et al إن درجة الحرارة المثلى لأنتاج إنزيم البروتيز من عزلة Aspergillus brasiliensis BCW2. عن طريق استعمال تخمرات الحالة الصلبة هي 30 مْ, ايضاً يختلف حجم اللقاح المستعمل بأختلاف نوع وسلالة الكائن المجهري فضلاً عن طبيعة عملية التخمر، اذ يعد وسيلة لقياس نمو الكائنات الحية الدقيقة وأمرًا حيويًا لتحقيق أقصى كمية من الناتج المرغوب , كما أن انخفاض حجم اللقاح المضاف قد لا يكون كافيًا لبدء نمو الاحياء المجهرية أو أنه يستغرق وقتًا طويلاً لنمو وتكاثر الاحياء المجهرية ، من ناحية أخرى ، بالمقابل يؤدي ارتفاع حجم اللقاح إلى تقييد حركة الاحياء المجهرية مع تثبيط نموها وتقليل إنتاجية التخمير (Othman et al . 2023). اذ استعمل (Chimbekujwo et al . ( 2020 1 مل من حجم اللقاح لأنتاج انزيم البروتيز من عزلة Aspergillus brasiliensis ، كما ذكر Bensmail et al. ( 2015) أن حجم اللقاح الأمثل لإنتاج انزيم البروتيز من عفن Aspergillus niger كان 1 × 106 بوغ / مل . كما ان هناك ارتباطاً وثيقاً مابين إنتاج الإنزيمات الميكروبية ومدة التحضين وهذا يعود لوجود علاقة مابين انتاج الانزيمات ونمو الخلايا ، أذ تًعد مدة الحضن من العوامل المهمة التي تؤثر على نمو وانتاج الأحياء المجهرية ، وفترة التحضين المثلى لنمو الكائن المجهري لأنتاج الانزيمات تتأثر بعدة عوامل منها مكونات الوسط الزرعي ونوع الكائن المجهري (Park et al . 1979). أذ تختلف فترة التحضين للحصول على أقصى إنتاجية للإنزيم بين الأنواع المختلفة أو حتى في نفس الأنواع المعزولة من مصادر مختلفة ، فقد اشار (Chimbekujwo et al. ( 2020 ان فترة التحضين المثلى لإنتاج انزيم البروتيز من عزلة Aspergillus brasiliensis BCW2 هي عند 72 ساعة ، كما ذكرUsman et al. (2021) إن أفضل مدة حضن لإنتاج إنزيم البروتيز ألحامضي من الفطريات الخيطية كانت 120 ساعة .
تنقية الإنزيم
وهي سلسلة متتابعة من الخطوات التي يمكن بها فصل الانزيم عن بقية المواد والبروتينات الاخرى الموجودة في المستخلص الخام والتي يمكن من خلالها ان تزداد الفعالية النوعية للأنزيم ، ولقد استخدمت عدة طرق لتنقية الانزيم (Banerjee et al., 2020) ، ومنها الترسيب بواسطة الاملاح مثل كبريتات الامونيوم والتي تعتمد على اساس معادلة الشحنات على سطح البروتين ، أذ تم اختيارها بسبب الذوبانية العالية لها في الماء حيث يزداد تركيز الملح في كل مرحلة لكي يتم ترسيب البروتينات الاخرى والتي يتم التخلص منها فيما بعد ، وبعد ذلك يذاب الانزيم بأستخدام دارىء معين او بالماء المقطر عن طريق النضح الغشائي بواسطة استخدام اكياس الديلزة ، ويمكن ترسيب الانزيمات ايضاً عن طريق استخدام المذيبات العضوية مثل الاسيتون والكحول الاثيلي من خلال خفض نقطة التعادل الكهربائي مما يؤدي الى زيادة قوة الجذب بين جزيئات البروتين وبالتالي ترسيبه (Fairooz et al., 2021)، والقسم الاخر من طرق التنقية والتي تعتمد على اساس الوزن الجزيئي مثل طريقة النضح الغشائي Dialysis وطريقة الترشيح الفائق Ultra Filtration وطريقة الترشيح الهلامي Gel filtration . والبعض الأخر منها يعتمد على مبدأ الشحنات مثل طريقة الترحيل الكهربائي Electrophoresis وطريقة كروموتوغرافيا التبادل الايوني Ion-exchange chromatography (Kamal et al. 2017). قام ( 2020 ) . Mamo et al بتنقية انزيم بروتيز الاسبارتك المنتج من عفن Aspergillus oryzae DRDFS13 تحت تخمرات الحالة الصلبة بواسطة تقنية كروماتوغرافيا التبادل الايوني (IEC) باستعمال عمود DEAE Sepharose fast flow بفعالية انزيمية (477.11 ) وحدة / مل وفعالية نوعية ( 183.50 ) وحدة / ملغم بعدد مرات تنقية 6.20 وبحصيلة انزيمية 9.2 % . كما تمكن ( 2014 ) . Hsiao et al بتنقية انزيم بروتيز الاسبارتك المنتج من عفن Rhizopus oryzae بواسطة تقنية AKTA Purifier 10 على عمود Q Sepharose بعدد مرات تنقية 22 وبحصيلة انزيمية 90.6 % ومن ثم استخدم عمود Phenyl Sepharose بعدد مرات تنقية 3.4 وبحصيلة انزيمية 58.8% .
توصيف إنزيم البروتيز
- الوزن الجزيئي
تختلف البروتيزات في اوزانها الجزيئية نتيجة لأختلاف المصادر المستخلصة منها ، اذ تستعمل عدة طرق لتحديد الاوزان الجزيئية مثل الترشيح الهلامي Gel-Filtration والترحيل الكهربائي Electrophoresis بوجود المادة الماسخة SDS والنبذ الفائق السرعة Ultracentrifuge ، ويعد الترحيل الكهربائي والترشيح الهلامي هي من اكثر الطرق المستخدمة لقياس الوزن الجزيئي Devi and Hemalatha ,2014) (.
أشار ( 2023 ) . Wei et al الى ان الوزن الجزيئي لأنزيم بروتيز الاسبارتك المنتج من عفن Aspergillus niger هو 50 كيلودالتون بأستعمال تقنية الترحيل الكهربائي Electrophoresis وبأستعمال SDS – PAGE . ولاحظ Ao et al .,(2018) ان الوزن الجزيئي لأنزيم بروتيز الاسبارتك المنتج من عفن Aspergillus oryzae Y1 هو 45 كيلودالتون بأستعمال تقنية الترحيل الكهربائي Electrophoresis . لاحظ ( 2020 ), Chimbekujwo et al ان الوزن الجزيئي لأنزيم بروتيز الاسبارتك المنتج من عفن Aspergillus brasiliensis BCW2هو 68 كيلودالتون بأستعمال تقنية الترحيل الكهربائي Electrophoresis وبوحود المادة الماسخة SDS – PAGE .
لاحظ ( 2023 ) . Tarek et al ان الوزن الجزيئي للبروتيز المستخلص من بكتريا Bacillus siamensis CSB55 هو 25 كيلودالتون بأستعمال تقنية الترحيل الكهربائي Electrophoresis . اشار ( 2021 ) . Karray et al الى ان الوزن الجزيئي للبروتيز المنتج من بكتريا Bacillus stearothermophilus هو 28 كيلودالتون بأستعمال تقنية الترحيل الكهربائي Electrophoresis . كما لاحظ Devi and Hemalatha . (2014) إن الوزن الجزيئي للبروتيز المستخلص من بذور نبات البطيخ هو 54 كيلودالتون بأستعمال طريقة الترشيح الهلامي Gel- Filtration وباستخدام عمود Sephadex G-200 . وأوضح . (2013) Al-Soufiإلى إن الوزن الجزيئي للبروتيز المستخلص من اوراق نبات الخس الشوكي Lactuca serriola L. هو 31000 دالتون بأستعمال طريقة الترشيح الهلامي Gel- Filtration وباستخدام عمود Sephacryl S-200 . لاحظ (2011) Yadav ان الوزن الجزيئي للبروتيز المستخلص من نبات Euphorbia neriifolia Linn 35.24 كيلودالتون بأستعمال تقنية الترحيل الكهربائي Electrophoresis وبوحود المادة الماسخة SDS – PAGE .
- تأثير الرقم الهيدروجيني على فعالية الإنزيم وثباته
تتأثر فعالية وثباتية الانزيم بالرقم الهيدروجيني ، وتكون الفعالية القصوى لكل انزيم عند رقم هيدروجيني معين يسمى بالرقم الهيدروجيني الامثل وتقل فعالية الانزيم عند الانحراف عن هذا الرقم (Kuddus., 2018 ) ، إذ تعتمد الأرقام الهيدروجينية المثلى للأنزيمات على عدة عوامل مثل طبيعة المحلول الدارىء والقوة الايونية ودرجة الحرارة والمادة الخاضعة وتركيز ومصدر الانزيم وتركيز المثبطات والمنشطات للأنزيم ، وقد يسبب التغير في الرقم الهيدروجيني الى حدوث مسخ Denaturation في تركيب البروتين مما يؤدي الى التأثير في طبيعة الانزيم وفعاليتة ، وللرقم الهيدروجيني تأثير على ايونية الاحماض الامينية الموجودة في الموقع الفعال للأنزيم مما يؤثر على الفعالية الإنزيمية الكلية نتيجة لتأين مكونات وسط التفاعل مثل تأين معقد الانزيم والمادة الخاضعة وتأين المادة الخاضعة فضلاً عن تأثيره الكبير في ثباتيه الانزيم (Segel,1976) . إذ أشار Mamo et al. (2020) ان ثبات فعالية الانزيم المنتج من عفنAspergillus oryzae DRDFS13 كانت في رقم هيدروجيني يتراوح بين ( 4.5- 6.5 ) في حين ان الرقم الهيدروجيني الامثل لأنزيم البروتيز كان 5 . وأكد ( 2021). Guo et al إن ثباتيه إنزيم البروتيز المنتج من عزلة Penicillium sp. XT7 كانت في رقم هيدروجيني يتراوح بين ( 3-6)، بينما أظهر أعلى نشاط له عند الرقم الهيدروجيني 3 .
وذكر ( 2021 ) . Karray et al ان ثبات فعالية أنزيم البروتيز المنتج من بكتريا Bacillus stearothermophilus كانت في رقم هيدروجيني يتراوح بين ( 7- 11 ) في حين ان الرقم الهيدروجيني الامثل كان 10 . واشار ( 2023 ) . Wei et al ان ثباتيه فعالية انزيم البروتيز المنتج من عفن Aspergillus niger في رقم هيدروجيني يتراوح بين ( 2- 5 ) والرقم الهيدروجيني الامثل للأنزيم كان 3 . وكما ذكر (Hsiao et al . (2014 ان ثباتية فعالية انزيم البروتيز المنتج من عفن Rhizopus oryzae كانت في رقم هيدروجيني يتراوح بين ( 3- 3.6 ) في حين إن الرقم الهيدروجيني الامثل للأنزيم كان (3.4) . واشار ( 2020 ) Chimbekujwo et al. إن ثباتية إنزيم البروتيز المنتج من عفن Aspergillus brasiliensis strain BCW2 كانت في رقم هيدروجيني يتراوح بين ( 4- 6 ) بينما كان الرقم الهيدروجيني الأمثل للأنزيم كان 9 .
- تأثير درجة الحرارة على فعالية الإنزيم وثباته
تؤثر درجة الحرارة في سرعة التفاعل الإنزيمي نتيجة لتأين المجاميع الفعالة لكل من الانزيم والمادة الخاضعة ، وتكون الفعالية القصوى لكل إنزيم عند درجة حرارة معينة تسمى بدرجة الحرارة المثلى ، ويعتمد تأثير درجة الحرارة في فعالية الإنزيم على تركيز المادة الخاضعة وثباتها وطبيعة بروتين الانزيم وثباته والالفة بين الانزيم والمادة الخاضعة والقوة الايونية ووجود المنشطات والمثبطات والرقم الهيدروجيني ، فضلاً عن ان وجود بعض المواد الأخرى مع الانزيم مثل الكاربوهيدرات والبروتينات التي تعطي حماية للأنزيم وتجعله اكثر مقاومة للحرارة ، فألانزيمات النقية لا تستطيع مقاومة الارتفاع في درجات الحرارة بينما الانزيمات غير النقية تكون اكثر مقاومة للحرارة ( Segel ,1976 ; Whitaker ,1972) .
اشار ( 2023 ) . Wei et al ان درجة الحرارة المثلى لأنزيم البروتيز المنتج من عفن Aspergillus niger كانت 50 م واحتفظ الإنزيم بنشاطه بنسبة 81% عند درجة حرارة 60 م ، لكن لم يظهر اي نشاط للأنزيم عند درجة حرارة 65 م وعندما تم تحضين الانزيم عند درجة حرارة 30- 50 م لمدة 3 ساعات، أظهر الإنزيم ثباتًا عاليًا، بينما انخفض نشاط الإنزيم بشكل ملحوظ إلى ما بعد 50 مْ ، وتم تعطيله تمامًا بعد 10 دقائق من التحضين عند 60 مْ . لاحظ ( 2021) . Guo et al ان درجة الحرارة المثلى لأنزيم البروتيز المنتج من عزلة Penicillium sp. XT7 كانت 30 درجة مئوية واحتفظ الانزيم بنسبة 86.8% من الحد الأقصى للنشاط عند 20 درجة مئوية وانخفض نشاطه المتبقي بشكل ملحوظ بعد الحضن عند 50 درجة مئوية. كما أشار (Souza et al. (2017 عند توصيف البروتيز الحامضي المنتج من عفن Aspergillus foetidus ان درجة الحرارة المثلى لأنزيم البروتيز هو (55 ) م وان ثباتية الانزيم تكون عند مدى واسع من درجات الحرارة تتراوح بين (30-60 ) م .
ولاحظ (Sampaio et al. (2011 عند دراسته تأثير درجة الحرارة على الأنزيم البروتيني الحامضي المنقى والمنتج من عفن Aspergillus clavatus عند مدى واسع من درجات الحرارة تراوحت بين (20-65 ) مْ ان درجة الحرارة المثلى لأنزيم البروتيز هو (55 ) م وان ثباتية الانزيم تكون عند مدى من درجات الحرارة تراوحت بين (30-60 ) م . اشار ( 2023 ) . Tarek et al ان درجة الحرارة المثلى لأنزيم البروتيز المنتج من عفن Bacillus siamensis CSB55 كانت 55 م وان ثباتية الانزيم تكون عند مدى من درجات الحرارة تراوحت بين (30-80 ) م واحتفظ الإنزيم بنشاطه بنسبة 80% عند درجة حرارة 40 و50 و60 و70 و80 مْ لمدة 16 و14 و10 و6 و4 ساعات على التوالي ، واحتفظ الإنزيم بنشاطه بنسبة 50% عند حضنه عند درجة حرارة 80 و 90 مْ لمدة 4 و2 ساعة على التوالي .
- تأثير الثوابت الحركية
تعد الثوابت الحركية والتي تشمل السرعة القصوى (Vmax) وثابت ميكالس(Km) من القيم المهمة للتفاعلات الإنزيمية ، ، اذ يعرف ثايت ميكالس (Km) بأنه تركيز المادة الخاضعة التي تكون عندها سرعة التفاعل الانزيمي تساوي نصف السرعة القصوى ، وتعبر قيمة ثابت ميكالس (Km) عن الألفة بين الانزيم والمادة الخاضعة ، اذ ان الفعالية الإنزيمية تكون عالية للأنزيمات التي تكون فيها قيمة ثابت ميكالس واطئة وعلى العكس من ذلك تكون الفعالية الانزيمية واطئة للأنزيمات التي تكون فيها قيمة ثابت ميكالس عالية .,2000) Copeland ) .
بين Purushothaman et al . (2019) من خلال احتواء خليط التفاعل على تراكيز مختلفة من المادة الخاضعة (الهيموكلوبين ) تراوحت بين ( 0- 2 % ) وبأستعمال طريقة Lineweaver- Burk plot. ، ان قيم الثوابت الحركية (Km) و) Vmax (لأنزيم بروتيز الاسبارتك المنتج من عفن Aspergillus niger كانت 6.3 ملغم / مل (98.52 مايكرومتر) ، و 50 مايكرومول / دقيقة على التوالي عند رقم هيدروجيني 4 ودرجة حرارة 60 مْ . ولاحظ Ao et al. (2018) عند استعمال تراكيز مختلفة من المادة الخاضعة ( الكازين ) تراوحت بين ( 0.5 – 3 ) غرام / لتر عند رقم هيدروجيني 7 ودرجة حرارة 55 مْ وباستخدام طريقة Lineweaver- Burk. ، ان قيم الثوابت الحركية Km و Vmax لأنزيم البروتيز المنتج من عفن Aspergillus oryzae Y1 كانت 20.0769 ملغم /مل و 256.4103 غرام / مل في الدقيقة على التوالي . واشار ( 2023 ) . Wei et al عند استعمال تراكيز مختلفة من المادة الخاضعة ( الكازين ) تراوحت بين ( 0.2 – 3 ) غرام / لتر عند رقم هيدروجيني 3 ودرجة حرارة 40 مْ وباستخدام طريقة Lineweaver- Burk. ، ان قيم الثوابت الحركية (Km) و) Vmax (لأنزيم البروتيز المنتج من عفن Aspergillus niger كانت 2.44 ملغم /مل و 83.54 وحدة / ملغم على التوالي . كما استطاع ( 2015 ) . Niyonzima et al من تحديد قيمة الثوابت الحركية (Km) و) Vmax (لأنزيم البروتيز المنتج من عفن Aspergillus terreus gr وكانت 5.4 ملغم /مل و 12.8 وحدة لكل مل على التوالي عند رقم هيدروجيني 11 ودرجة حرارة 50 مْ وباستخدام طريقة Lineweaver- Burk. .
- تأثير العناصر المعدنية والمنشطات والمثبطات على فعالية انزيم بروتيز الاسبارتك
تؤثر المركبات والايونات المعدنية على فعالية الإنزيمات ، فبعضها يرتبط بالإنزيم ويزيد من فعاليته وتسمى بالمنشطات Activators والبعض الاخر يقلل من فعالية الانزيم وتسمى بالمثبطات Inhibitor ، (Usharani et al., 2010). ان تأثير المثبطات على المادة الخاضعة يكون من خلال ارتباطها بالموقع الفعال مما يؤدي الى تقليل الالفة بين المادة الخاضعة والانزيم ، اذ ان عملية تثبيط الانزيم تعتمد على تركيز المثبط وتركيز المادة الخاضعة وطبيعتها وتركيز الإنزيم ومصدره ووقت التفاعل ودرجة الحرارة والرقم الهيدروجيني والقوة الأيونية Panesar et al.,2006) ( .
أشار ( 2020 ) . Song et al الى ان ايونات Cu2+ و Mn2+ كان لها تأثير في زيادة فعالية انزيم البروتيز المنتج من عفن Aspergillus niger F0215 في حين ان استعمال الببستاتين ادى الى تثبيط الانزيم بصورة كاملة . ذكر ( 2014 ) . Devi and HemaLatha ان انزيم البروتيز المنتج من بذور Cucumis melo Var agrestis قد ثُبط بصورة كاملة عند حضنه مع 5 ملي مولاري من مادة PMSF بينما تأثر الانزيم قليلاً عند حضنه مع مادة EDTA .
وفي دراسة قام بها Marcial et al. (2011) لأنتاج انزيم بروتيز الاسبارتك من فطر Amylomyces rouxii ، اذ اظهرت النتائج ان استعمال 3 ملي مولاري من الببستاتين يعمل على تثبيط الانزيم بنسبة 73% ، الا ان استعمال 1 ملي مولاري من PMSF يعمل على تثبيط عمل الانزيم بنسبة 12% ، في حين ان استعمال 1 ملي مولاري من EDTA يعمل على تثبيط عمل الانزيم بنسبة 20% . كما لاحظ (Hsiao et al. (2014 عدم وجود إي تأثير تثبيطي عند إضافة 10 ملي مولاري من EDTA لأنزيم البروتيز المنتج من عفن Rhizopus oryzae عند الرقم الهيدروجيني 3.4 ودرجة حرارة 35 مْ ، بينما حصل تثبيط عمل الانزيم بشكل كامل عند استعمال مادة الببستتاتين Pepstatin A ، في حين لم يكن لإضافة 1 ملي مولار Ca2+ أي تأثير على نشاط التحلل البروتيني، اما عند إضافة 1 ملي مولاري Zn2+ وNi2+ و Cu2+وMn2+ وCo2+ أدت إلى تقليل نشاط الإنزيم بشكل طفيف .
الخاتمة
يتزايد الطلب على الإنزيمات في القطاعات الصناعية بسرعة بسبب مزاياها الاقتصادية والبيئية. أذ تنتج الكائنات الحية الدقيقة أنواعًا مختلفة من الإنزيمات ومن بين هذه الانزيمات هي انزيمات البروتيزات المنتجة من مصادر ميكروبية نتيجة لأنخفاض التكلفة وارتفاع معدل الإنتاج والتي اكتسبت اهتمامًا خاصًا في القطاعات الصناعية وتعد الفطريات هي مصادر جيدة لأنتاج أنزيمات البروتيز وخاصة بروتيز الاسبارتك الذي ينتج من العزلات الفطرية التي تنتمي إلى جنس Aspergillus .
المصادر
- Sharma, N. (2019). A review on fungal alkaline protease. J. Emerg. Technol. Innov. Res. 6, 261–273. doi: 10.1729/Journal.22354
- Gimenes, N. C., Silveira, E., & Tambourgi, E. B. (2021). An overview of proteases: production, downstream processes and industrial applications. Separation & Purification Reviews, 50(3), 223-243.
- Pawar, K. S., Singh, P. N., & Singh, S. K. (2023). Fungal alkaline proteases and their potential applications in different industries. Frontiers in Microbiology, 14, 1138401
- Choudhary, K., Mankar, M. K., and Sahay, S. (2022). “Extremophilic enzymes: catalytic features and industrial applications” in Extremophilic fungi: Ecology, Physiology and Applications (Singapore: Springer Nature Singapore), 273–314.
- Koistinen, H., Kovanen, R. M., Hollenberg, M. D., Dufour, A., Radisky, E. S., Stenman, U. H., &Mirtti, T. (2023). The roles of proteases in prostate cancer. IUBMB life.
- Kalaiselvan, P., Dutta, D., Konda, N. V., Sharma, S., Kumar, N., Stapleton, F., & Willcox, M. D. (2023). Effect of Deposition and Protease Digestion on the Ex Vivo Activity of Antimicrobial Peptide-Coated Contact Lenses. Nanomaterials, 13(2), 349.
- Couture, F. (2023). Therapeutic Targeting of the Proteolytic Enzymes. International Journal of Molecular Sciences, 24(1), 521.
- Dasari, P. K., Gannamaneni, S. S., Jahnavi, B., Umadevi, B., Balaji, M. A., Meghana, N., &Sravani, R. (2023). optimization and production of protease using aspergillus cervinus.World Journal of Pharmaceutical Research , 12(4) , 2063-2071.
- Dudani, J. S., Warren, A. D., & Bhatia, S. N. (2018). Harnessing protease activity to improve cancer care. Annual review of cancer biology, 2, 353-376.
- Zhang, J., Qiao, W., & Luo, Y. (2023). Mitochondrial quality control proteases and their modulation for cancer therapy. Medicinal Research Reviews, 43(2), 399-436.
- Erekat, N. S. (2022). Programmed cell death in cerebellar Purkinje neurons. Journal of integrative neuroscience, 21(1), 30,1-15.
- Naveed, M., Nadeem, F., Mehmood, T., Bilal, M., Anwar, Z., & Amjad, F. (2021). Protease—a versatile and ecofriendly biocatalyst with multi-industrial applications: an updated review. Catalysis Letters, 151, 307-323.
- Abd AL-MAnhel, A. J. (2017). Production of exopolysaccharide from local fungal isolate. Current Research in Nutrition and Food Science Journal, 5(3), 338-46.
- Patil, P. M.;Kulkarni A. A. and Kininge , P.(2012). Production of Milk Clotting Enzyme from Aspergillus oryzae under Solid-State Fermentation using Mixture of Wheat Bran and Rice Bran. International Journal of Scientific and Research Publications,2(10):1-12.
- Mienda, B. S., Yahya, A., Galadima, I. A., & Shamsir, M. S. (2014). An overview of microbial proteases for industrial applications. Res J Pharm Biol Chem Sci, 5(1), 388-396.
- Mamo,J.; Assefa,F.(2018). The Role of Microbial Aspartic Protease Enzyme in Food and Beverage Industries. Journal of Food Quality,2018:1-16.
- Rawlings, N. D., & Salvesen, G. (2013). Handbook of proteolytic enzymes (No. 8040). Academic press.
- Gurumallesh, P., Alagu, K., Ramakrishnan, B., & Muthusamy, S. (2019). A systematic reconsideration on proteases. International journal of biological macromolecules, 128, 254-267.
- majid dakhil AL-Sefar, R. (2017). Purification and Charactrization of protease for some plants sources. Al-Qadisiyah Journal of Pure Science, 22(4), 105-129.
- Gimenes, N. C., Silveira, E., & Tambourgi, E. B. (2021). An overview of proteases: production, downstream processes and industrial applications. Separation & Purification Reviews, 50(3), 223-243.
- Wei, M., Chen, P., Zheng, P., Tao, X., Yu, X., & Wu, D. (2023). Purification and characterization of aspartic protease from Aspergillus niger and its efficient hydrolysis applications in soy protein degradation. Microbial Cell Factories, 22(1), 42,1-9 .
- Mamo,J.; Orellana,J.F.S.; Yeleman, V.; Kangwa,M.; Fernandez-Lahore,M.H and Assefa,F(2020). Purification and Characterization of Aspartic Protease Produced from Aspergillus oryzae DRDFS13 under Solid-State Fermentation. bioRxiv, 2020,1-38
- Singh, R., Mittal, A., Kumar, M., & Mehta, P. K. (2016). Microbial proteases in commercial applications. J Pharm Chem Biol Sci, 4(3), 365-374.
- Razzaq, A., Shamsi, S., Ali, A., Ali, Q., Sajjad, M., Malik, A., & Ashraf, M. (2019). Microbial proteases applications. Frontiers in bioengineering and biotechnology, 7, 110,1-20 .
- Banerjee, G., & Ray, A. K. (2017). Impact of microbial proteases on biotechnological industries. Biotechnology and Genetic Engineering Reviews, 33(2), 119-143.
- Solanki, M. K., Abdelfattah, A., Sadhasivam, S., Zakin, V., Wisniewski, M., Droby, S., & Sionov, E. (2021). Analysis of stored wheat grain-associated microbiota reveals biocontrol activity among microorganisms against mycotoxigenic fungi. Journal of Fungi, 7(9), 781, 1-16.
- Machado, A. R. G., Teixeira, M. F. S., de Souza Kirsch, L., Campelo, M. D. C. L., & de Aguiar Oliveira, I. M. (2016). Nutritional value and proteases of Lentinus citrinus produced by solid state fermentation of lignocellulosic waste from tropical region. Saudi Journal of Biological Sciences, 23 (5), 621-627.
- Mandujano-González, V., Villa-Tanaca, L., Anducho-Reyes, M. A., & Mercado-Flores, Y. (2016). Secreted fungal aspartic proteases: A review. Revista Iberoamericana de Micología, 33(2), 76-82.
- Da Silva, R. R. (2017). Bacterial and fungal proteolytic enzymes: production, catalysis and potential applications. Applied biochemistry and biotechnology, 183, 1-19.
- Souza, P. M., Werneck, G., Aliakbarian, B., Siqueira, F., Ferreira Filho, E. X., Perego, P., … & Junior, A. P. (2017). Production, purification and characterization of an aspartic protease from Aspergillus foetidus. Food and Chemical Toxicology, 109, 1103-1110.
- Contesini, F. J., Liberato, M. V., Rubio, M. V., Calzado, F., Zubieta, M. P., Riaño-Pachón, D. M., & Damasio, A. R. (2017). Structural and functional characterization of a highly secreted α-L-arabinofuranosidase (GH62) from Aspergillus nidulans grown on sugarcane bagasse. Biochimica et Biophysica Acta (BBA)-proteins and proteomics, 1865(12), 1758-1769.
- Ao, X. L., Yu, X., Wu, D. T., Li, C., Zhang, T., Liu, S. L., Chen, S.J., He ,L., Zhou, K., Zou, L. K. (2018). Purification and characterization of neutral protease from Aspergillus oryzae Y1 isolated from naturally fermented broad beans. AMB Express, 8(1), 1-10.
- De Oliveira, D. M., Forde, B. M., Kidd, T. J., Harris, P. N., Schembri, M. A., Beatson, S. A., Paterson, D. L., Walker, M. J. (2020). Antimicrobial resistance in ESKAPE pathogens. Clinical microbiology reviews, 33(3), 10-1128.
- Zhang, B., Sun, Q., Liu, H. J., Li, S. Z., & Jiang, Z. Q. (2017). Characterization of actinidin from Chinese kiwifruit cultivars and its applications in meat tenderization and production of angiotensin I-converting enzyme (ACE) inhibitory peptides. LWT, 78, 1-7.
- Mothe, T., & Sultanpuram, V. R. (2016). Production, purification and characterization of a thermotolerant alkaline serine protease from a novel species Bacillus caseinilyticus. 3. Biotech, 6(1), 53.
- Theron, L. W., & Divol, B. (2014). Microbial aspartic proteases: current and potential applications in industry. Applied microbiology and biotechnology, 98, 8853-8868.
- Brik, A., & Wong, C. H. (2003). HIV-1 protease: mechanism and drug discovery. Organic & biomolecular chemistry, 1(1), 5-14.
- Coates, L., Tuan, H. F., Tomanicek, S., Kovalevsky, A., Mustyakimov, M., Erskine, P., & Cooper, J. (2008). The catalytic mechanism of an aspartic proteinase explored with neutron and X-ray diffraction. Journal of the American Chemical Society, 130(23), 7235-7237.
- Wångsell, F. (2006). Design and Synthesis of Serine and Aspartic Protease Inhibitors (Doctoral dissertation, Institutionen för fysik, kemi och biologi).
- Naveed, M., Nadeem, F., Mehmood, T., Bilal, M., Anwar, Z., & Amjad, F. (2021). Protease—a versatile and ecofriendly biocatalyst with multi-industrial applications: an updated review. Catalysis Letters, 151, 307-323.
- N Jisha, V., B Smitha, R., Pradeep, S., Sreedevi, S., N Unni, K., Sajith, S., … & Benjamin, S. (2013). Versatility of microbial proteases. Advances in enzyme research, 1(03), 39-51.
- Dadshahi, Z., Homaei, A., Zeinali, F., Sajedi, R. H., & Khajeh, K. (2016). Extraction and purification of a highly thermostable alkaline caseinolytic protease from wastes Penaeus vannamei suitable for food and detergent industries. Food Chemistry, 202, 110-115.
- Sun,Q.;Chen,F.;Geng,F.;Luo,Y.;Gong,S. and Jiang,Z.(2018).A novel aspartic protease from Rhizomucor miehei expressed in Pichia pastoris and its application on meat tenderization and preparation of turtle peptides. Food Chemistry,245:570-577.
- Putatunda, C., Kundu, BS., Bhatia, R . (2019) Purification and characterization of alkaline protease from Bacillus sp. HD292. Proc Natl Acad Sci India B Biol Sci 89:957–965. https:// doi. org/ 10. 1007/ s40011- 018- 1011-z
- Bhatia, RK., Ullah, S., Hoque, HZ., Ahmad, I., Yang, YH., Bhatt, AK., Bhatia, SK .(2021) Psychrophiles: a source of cold-adapted enzymes for energy efficient biotechnological industrial processes. J Envir Chem Eng 9:104607. https:// doi. org/ 10. 1016/j. jece. 2020. 104607
- Jadhav, H. P., Sonawane, M. S., Khairnar, M. H., and Sayyed, R. Z. (2020) Production of alkaline protease by rhizospheric Bacillus cereus HP_RZ17 and Paenibacillus xylanilyticus HP_RZ19. Environ Sustain 3:5–13. https:// doi. org/ 10. 1007/ s42398- 020- 00096-z
- Hakim, A., Bhuiyan, F. R., Iqbal, A., Emon, T. H., Ahmed, J., & Azad, A. K. (2018) Production and partial characterization of dehairing alkaline protease from Bacillus subtilis AKAL7 and Exiguobacterium indicum AKAL11 by using organic municipal solid wastes. Heliyon 4:e00646. https:// doi. org/ 10. 1016/j. heliy on. 2018. e00646 .
- Ash, K., Sushma, Ramteke, PW. (2018) Optimization of extracellular alkaline protease production from Pseudomonas aeruginosa isolated from soil samples. Inter J Agric Environ Biotechnol 11:187–194. https:// doi. org/ 10. 30954/ 0974- 1712. 2018. 00178. 24
- Prajapati, V. S., Ray, S., Narayan, J., Joshi, C. C., Patel, K. C., Trivedi, U. B., & Patel, R. M. (2017) Draft genome sequence of a thermostable, alkaliphilic α -amylase and protease producing Bacillus amyloliquefaciens strain KCP2. 3 Biotech (2017) 7:372 .
- Usman, A., Mohammed, S., &Mamo, J. (2021). Production, optimization, and characterization of an acid protease from a filamentous fungus by solid-state fermentation. International journal of microbiology, 2021.
- Novelli, P.K., Barros, M.M., Fleuri, L.F., (2016). Novel inexpensive fungiproteases: production by solid state fermentation and characterization. Food Chem. 198,119e124.
- Yin, F. W., Sun, X. L., Zheng, W. L., Yin, L. F., Luo, X., Zhang, Y. Y., Wang,Y.F., Fu, Y. Q. (2023). Development of a Strategy for L-Lactic Acid Production by Rhizopus oryzae Using Zizania latifolia Waste and Cane Molasses as Carbon Sources. Molecules, 28(17), 6234.
- Patil, P. M.;Kulkarni A. A. and Kininge , P.(2012). Production of Milk Clotting Enzyme from Aspergillus oryzae under Solid-State Fermentation using Mixture of Wheat Bran and Rice Bran. International Journal of Scientific and Research Publications,2(10):1-12.
- Daudi,S.; Mukhtar,H.; Rehman,A.U. and UL-Haq, I.(2015). Production of rennin-like acid protease by Mucor pusillus through submerged fermentation .Pak.J.Bot.,47(3):1121-1127.
- Aljammas, H. A., Al Fathi, H., & Alkhalaf, W. (2018). Study the influence of culture conditions on rennin production by Rhizomucor miehei using solid-state fermentations. Journal of Genetic Engineering and Biotechnology, 16(1), 213-216.
- Purushothaman, K., Bhat, S. K., Singh, S. A., Marathe, G. K., & Rao, A. R. G. A. (2019). Aspartic protease from Aspergillus niger: Molecular characterization and interaction with pepstatin A. International journal of biological macromolecules, 139, 199-212.
- Yujie, G.; Tao, T.; Peng, Y.; Yaru, W. ; Yaxin, R. ; Bin, Y. and L. Huiying, (2019).High-level expressionand characterization of a novel aspartic protease from Talaromyces leycettanus JCM12802 and its potential application in juice clarification, Food Chem. 281: 197–203.
- dos Santos Aguilar, J. G., & Sato, H. H. (2018). Microbial proteases: Production and application in obtaining protein hydrolysates. Food Research International, 103, 253-262.
- Othman, B., & Sebo, N. H. (2023). Utilization of Some Agro Wests for the Production of Acid Protease by Aspergillus niger. Journal of Survey in Fisheries Sciences, 10(3S), 4319-4331.
- Hamza, T. A., & Woldesenbet, F. (2017). Optimization of culture growth parameters for production of protease from bacteria, isolated from soil. Biosci Bioeng, 3, 1-0.
- Aunstrup, K., Andersen, O., Falch, E. A. and Nielsen, T.K. ( 1979 ). Production of microbial enzymes, In Microbiol Technology, A.H. Rose, ed. Academic press, Inc, New York .
- Bano, S., Dahot, M. U., & Naqvi, S. H. A. (2016). Optimization of culture conditions for the production of protease by Pleurotus eryngii. Pakistan Journal of Biotechnology, 13(3), 193-198.
- Bensmail, S., Mechakra, A., & Fazouane-Naimi, F. (2015). Optimization of milk-clotting protease production by a local isolate of Aspergillus niger ffb1 in solid-state fermentation. The journal of microbiology, biotechnology and food sciences, 4(5), 467.
- Luti, K., &Younis, R. (2014). An induction of undecylprodigiosin production from Streptomyces coelicolor by elicitation with microbial cells using solid state fermentation. Iraqi J. Sci, 55, 553-1562.
- Chahrazad, B., & Habiba, Z. (2021). Production and characterization of proteases from Aspergillus niger strain isolated from dried fruit and its effect on gliadin. Algerian Journal of Nutrition and Food Sciences, 1(4), 25–29
- Khademi, F., Abachi, S., Mortazavi, A., Ehsani, M. A., Tabatabaei, M. R., & Malekzadeh, F. A. (2013). Optimization of fungal rennet production by local isolate of Rhizomucor nainitalensis under solid substrate fermentation system. IOSR Journal of Pharmacy and Biological Sciences, 5(2), 115-121.
- Foda, M. S., Moharam, M. E., Ramadan, A., & El-Bendary, M. A. (2012). Over production of milk clotting enzyme from Rhizomucor miehei through adjustment of growth under solid state fermentation conditions. Australian Journal of Basic and Applied Sciences, 6(8), 579-589.
- Chimbekujwo, K. I., Ja’afaru, M. I., & Adeyemo, O. M. (2020). Purification, characterization and optimization conditions of protease produced by Aspergillus brasiliensis strain BCW2. Scientific African 8: e00398.
- Park, Y. K., De Santi, M. S. S., & Pastore, G. M. (1979). Production and characterization of β‐galactosidase from Aspergillus oryzae. Journal of Food Science, 44(1), 100-103.
- Fairooz, H. A., Fairooz, N. A., & Chaloob, F. A. (2021). Extraction condition of protease frombroccoli (Brassicaoleracea var italic) and study of Biochemical Characteristic fromthe selectedsource. Annals of the Romanian Society for Cell Biology, 2208-2219.
- Kamal, S., Rehman, S., & Iqbal, H. M. (2017). Biotechnological valorization of proteases: from hyperproduction to industrial exploitation—a review. Environmental Progress & Sustainable Energy, 36(2), 511-522.
- Hsiao, N. W., Chen, Y., Kuan, Y. C., Lee, Y. C., Lee, S. K., Chan, H. H., & Kao, C. H. (2014). Purification and characterization of an aspartic protease from the Rhizopus oryzae protease extract, Peptidase R. Electronic Journal of Biotechnology, 17(2), 89-94.
- Devi, B. G., & HemaLatha, K. P. J. (2014). Isolation, partial purification and characterization of alkaline serine protease from seeds of Cucumis melo var agrestis. International Journal of Research in Engineering and Technology, 3(6), 88-95.
- Tarek, H., Nam, K. B., Kim, Y. K., Suchi, S. A., & Yoo, J. C. (2023). Biochemical Characterization and Application of a Detergent Stable, Antimicrobial and Antibiofilm Potential Protease from Bacillus siamensis. International Journal of Molecular Sciences, 24(6), 5774.
- Karray, A., Alonazi, M., Horchani, H., & Ben Bacha, A. (2021). A novel thermostable and alkaline protease produced from Bacillus stearothermophilus isolated from olive oil mill sols suitable to industrial biotechnology. Molecules, 26(4), 1139.
- Al-Soufi, M. A. (2013). Partial purification and estimated some characterization of protease from prickly lettuce (Lactuca serriola L.) leaves and used it in some applications. Iraqi Journal of Biotechnology, 12(2), 1-18.
- Yadav, R. P., Patel, A. K., & Jagannadham, M. V. (2011). Purification and biochemical characterization of a chymotrypsin-like serine protease from Euphorbia neriifolia Linn. Process Biochemistry, 46(8), 1654-1662.
- Kuddus, M. (Ed.). (2018). Enzymes in food biotechnology: production, applications, and future prospects.912 .
- Guo, Y., Li, X., Jia, W., Huang, F., Liu, Y., & Zhang, C. (2021). Characterization of an intracellular aspartic protease (PsAPA) from Penicillium sp. XT7 and its application in collagen extraction. Food Chemistry, 345, 128834.
- Sampaio e Silva, T. A., Knob, A., Tremacoldi, C. R., Brochetto-Braga, M. R., & Carmona, E. C. (2011). Purification and some properties of an extracellular acid protease from Aspergillus clavatus. World Journal of Microbiology and Biotechnology, 27, 2491-2497.
- Copeland, R. A. (2000). Kinetics of single-substrate enzyme reactions. Enzymes: A Practical Introduction to Structure, Mechanism, and Data Analysis, 2, 137-139.
- Niyonzima, F. N., & More, S. S. (2015). Purification and characterization of detergent-compatible protease from Aspergillus terreus gr. 3 Biotech, 5, 61-70.
- Usharani, B., & Muthuraj, M. (2010). Production and characterization of protease enzyme from Bacillus laterosporus. Afr. J. Microbiol. Res, 4(11), 1057-1063.
- Panesar, P. S., Panesar, R., Singh, R. S., Kennedy, J. F., & Kumar, H. (2006). Microbial production, immobilization and applications of β‐D‐galactosidase. Journal of Chemical Technology & Biotechnology: International Research in Process, Environmental & Clean Technology, 81(4), 530-543.
- Song, P., Cheng, L., Tian, K., Zhang, M., Mchunu, N. P., Niu, D., … & Wang, Z. X. (2020). Biochemical characterization of two new Aspergillus niger aspartic proteases. 3 Biotech, 10, 1-9.
- Marcial, J., Pérez De Los Santos, A. I., Fernández, F. J., Díaz-Godínez, G., Montiel-González, A. M., & Tomasini, A. (2011). Characterization of an aspartic protease produced by Amylomyces rouxii. Revista mexicana de ingeniería química, 10(1), 9-16.